

Architecture has frequently drawn inspiration from nature in its forms and structures. More recently however, the inner logic of nature’s morphological processes is being appropriated by architects to create genetic algorithms. These algorithms, which were initially developed to aid biologists in understanding the dynamics of evolutionary processes, are being used by architects to guide the evolution of forms. This paradigm shift from form-making to form-evolving potentially limits the role of the designer to a kind of prize-dog breeder. This essay explores the new developments of Evo Devo in order to propose that by modifying the genetic algorithm to encompass Hox logic, architecture can progress into a period of ‘post-Cambrian’ architectural diversity.

Figure 1 (Cover): An evolutionary architecture by John Frazer (Frazer, 1995)
Figure 2 (Above) : Butterfly (http://upload.wikimedia.org/wikipedia/commons/6/65/Blue_morpho_butterfly.jpg, 25/4/09)

Figure 3 (Left): Fallingwater,1934 by Frank Lloyd Wright (http://en.wikipedia.org/wiki/Falling_water, 25/4/09)
Figure 4 (Right): Casa Milà, 1910 by Antonio Gaudí (Giralt-Miracle, 2002, p. 155)
Introduction

Figure 5: Butterfly curve (Available: http://local.wasp.uwa.edu.au/~pbourke/geometry/butterfly/, 25/4/09)
Architecture has recently undergone a paradigm shift from being based on purely visual concerns towards an architecture justified by its performance. This shift privileges material performance over appearance and process over representation (Leach, 2009, p. 34). The resulting design process can be described as an interest in morphogenesis. Derived from the Greek terms ‘morphe’ (shape/form) and genesis (creation), morphogenesis was initially used in the realm of biological sciences to refer to ‘the logic of form generation and pattern-making in an organism through processes of growth and differentiation’ (Leach, 2009, p. 34).
Nature as a form of inspiration is not something new to architecture – one only has to look at the works of Frank Lloyd Wright or Antonio Gaudí to see an attempt to harmonise the natural and the built world. These organic architects drew inspiration from nature’s forms and structures (Frazer, 1995, p. 10). Indeed, as the engineer Fred Severud claims ‘…it is a fact that the contemporary architect or engineer faces few problems in structural design which Nature has not already met and solved’ (Severud in Steadman, 2008, p. 261). More recently however, inspiration is being drawn from new developments in biology which are illuminating the inner logic of nature’s morphological processes. Biomimetics, the study of what we can learn by replicating the mechanisms of nature, has therefore emerged as an important field of research and is having particular resonance in morphogenetic architecture (Leach, 2009, p. 35).
When studying natural systems, scientists construct an algorithm which becomes a model of the system they are studying. These models are simplified, abstract descriptions of what they think is happening in a particular phenomenon (Ball, 2009, p. 30). The algorithms do not need to capture reality fully, just those aspects which are essential to the phenomenon. For this reason, much skill lies is deciding which information to include and which to exclude. The maxim, ‘a problem well put is a problem half-solved’ therefore becomes particular pertinent.
The key difference between the algorithms generated by scientists and those generate by architects is the role of the algorithm. Scientific algorithms are a model of a system; architectural algorithms are a model for a system. The architectural algorithm therefore must be able to go beyond mimicry and posses an inherent logic relevant to architecture. This paper explores the role of genetic algorithms in the architectural design process by focusing on how these algorithms can be modified to aid in the genesis of architectural form.
Algorithmic architecture

Figure 6: 3D cellular automata urbanism by Team Jungle, AADRL (Verebes, 2008, p. 77)
In its most general sense, an algorithm ‘is a process of addressing a problem in a finite number of steps. It can be an articulation of either ‘a strategic plan for solving a known problem or a stochastic search towards possible solutions to a partially known problem’ (Terzidis, 2006, p. 15). Algorithms are expressed in terms of mathematical equations which define the rules of the model. Nowadays, the term ‘algorithm’ has been extended to encompass both the rule for calculating something and the program for computing it (Gell-Mann, 1994, p. 35).
However, contrary to its digital connotations, it could be argued that algorithms have been used implicitly in architecture for centuries prior to the digital age. Terzidis (2006, p. 39) postulates that while the algorithm is often associated with computer science, the use of instructions, commands or rules in architecture are, in essence, algorithms. Hersey and Freedam (1992) for instance, were able to ‘detect, extract, and formulate rigorous geometric rules’ (Terzidis, 2006, p. 21) by which Palladio conceived his villas. It is therefore important to make explicit that when referring to ‘algorithm’, this paper is referring to it in the modern sense of the word to means both the algorithm and the program computing it.
Although, the etymology of ‘algorithm’ dates back to the 9th-century, explicit algorithmic architecture has emerged only in the past two decades. To date, only three main categories of algorithms are prominently used in the genesis of architectural form. These categories can be defined as follows: Fractals, which include self-similar and fractal geometry such as L-systems; Chaos, which refers to nonlinear dynamics and strange attractors such as the Lorenz Attractor; and finally, Complex Adaptive Systems, which deal with autonomous, self-organising agents as demonstrated in Cellular Automata or the Genetic Algorithm.
Morphogenetic architects have taken a particular affinity to Complex Adaptive Systems in large part to their principles of emergence and self-organization. Coined by John Holland and Gell-mann of the Santa Fe Institute, Complex Adaptive Systems depend on ‘extensive interactions, the aggregation of diverse elements, and adaption or learning’ (Holland, 1996, p. 4). This can be shown in the collective intelligence of biological systems such as a colony of ants or termites, slime mould, flocking birds or a school of fish. It can also be demonstrated in non-biological systems such as, the internet, the stock market or even cities. What becomes clear in these complex adaptive systems is that complex patterns emerge from remarkably few ingredients. So although unpredictable outcomes may emerge, the results are intrinsically connected through the rules that govern them.
In the early 1990s, ecologist Thomas Ray of the Sante Fe Institute took the notion of Complex Adaptive Systems one step further by introducing biological evolution into the simulation. His much acclaimed TIERRA program introduced the notion of a Genetic Algorithm which could simulate ‘the behavior and adaption of a population of candidate solutions over time as generations are created, tested, and selected through repetitive mating and mutation’ (Terzidis, 2006, p. 19). In order for the genetic algorithm to simulate evolution, selection criteria needed to be established to stipulate which agents were to breed. This criterion was termed ‘fitness’ and as a result, how one defines fitness is critical to how a genetic algorithm works.

Figure 7: A simplified genetic algorithm (Flake, 1998, p. 344)
To date, most genetic algorithms have been used mainly for search and optimisation problems such as devising strategies to win at checkers (Gell-Mann, 1994, p. 310). In these cases, fitness is clearly defined. But if architects are to implement genetic algorithms in their design process, how does one define architectural fitness? Architecture must address multiple criteria. But can these criteria be combined, as they must be, to give a single composite fitness score? It would seem that defining a single architectural fitness criterion is the biggest obstacle for architects wanting to implement genetic algorithms into their work.
Steadman (2008, p. 255) proposes that multiple fitness criteria can be combined if the principle of Pareto optimality is introduced. Borrowed from classical economics, a solution is Pareto optimal if performance cannot be improved on any one criterion without worsening performance of others. This is indeed one way to address the problem but perhaps the notions of fitness, selection and evolution are not the best way forward. It is at this juncture that the new development in biology might offer a better way forward for architects.

Figure 8: Fractals – Romanesco broccoli (http://en.wikipedia.org/wiki/Fractal, 25/4/09)
Figure 9: Chaos – Lorenz attractor (http://en.wikipedia.org/wiki/Lorenz_attractor, 25/4/09)
Figure 10: Complex adaptive system – Flocking birds (Aranda & Lasch, 2006, p. 65)
Unfolding biology

Figure 11: Endless forms most beautiful (http://picasaweb.google.com/laurakoontz/Chicago#5088582514572741842, 25/4/09)
There is a grandeur in this view of life, with its several powers, having been originally breathed into a few forms or into one: and that whilst this planet has gone cycling on according to the fixed law of gravity, from so simple a beginning endless forms most beautiful and most wonderful have been, and are being, evolved.
(Darwin, 1859)
The genetic algorithm developed by Ray is modelled on Darwin’s notion of evolution. Up until the latter half of the 19th-century, it was believed by many that the earth and nature were a product of a Divine Architect (Ball, 2009, pp. 7-8). It was not until Charles Darwin’s acclaimed book, ‘The origin of species’ published in 1859 that this notion was challenged. Darwin proposed a theory of evolution based on random mutation and natural selection which explained how apparent ‘design’ might arise in nature without a designer. He argued that species of life have evolved over time from common ancestors through the process of natural selection. Ball (2009, p. 10) summarises:
Given enough time, small random changes in the forms of organisms could carry them towards those shapes that were best adapted to the demands that their environment made on them, because the struggle for survival weeded out those changes that made survival harder while conferring a reproductive advantage on those individuals who, by sheer luck, acquired a beneficial mutation.
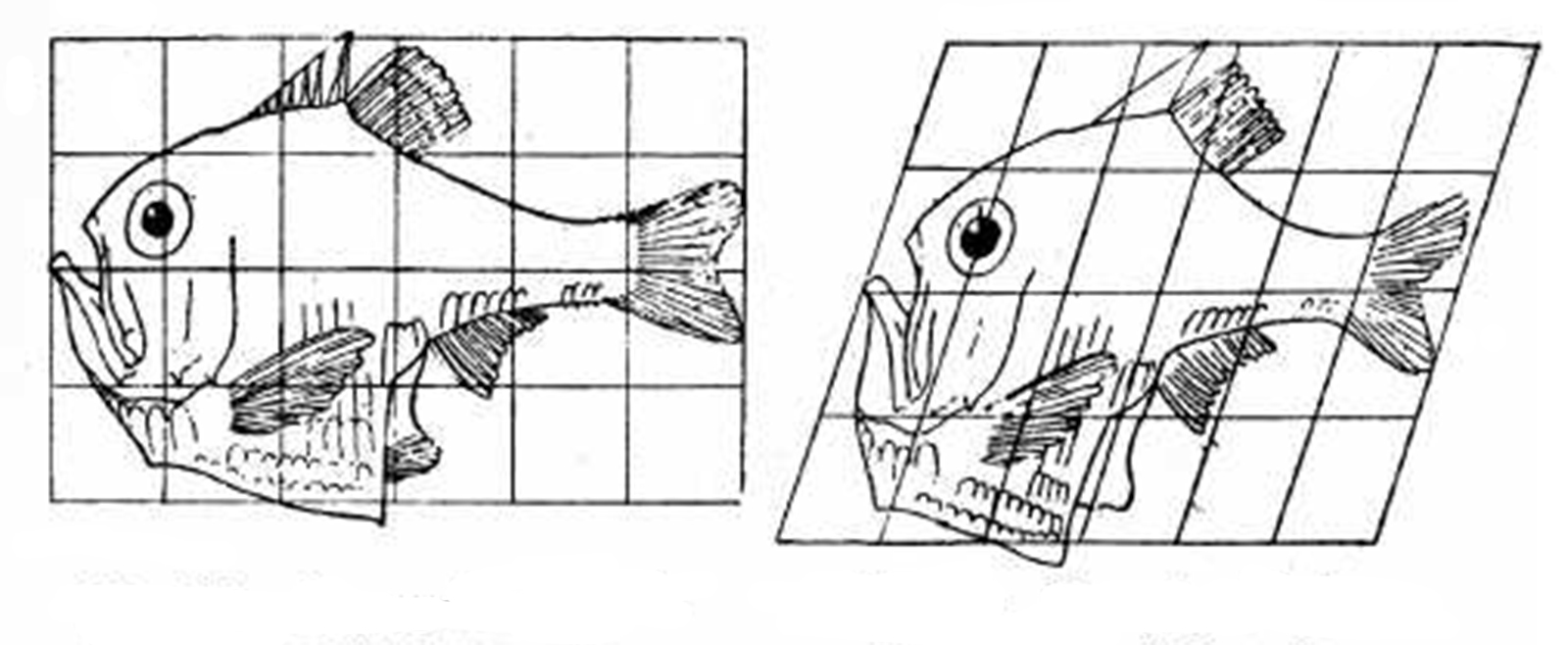
Figure 12: Transformation of Argyropelecus olfersi into Sternoptyx diaphana, Thompson (http://en.wikipedia.org/wiki/D’Arcy_Wentworth_Thompson, 25/4/09)
As Flake (1998, p. 339) claims of Darwin’s work, ‘never have so many natural phenomena been explained by so few facts.’ However, the notion of Darwinian evolution was not entirely satisfying in that it was essentially a narrative, not an explanation. It said nothing about mechanism, or in other words, it was a narrative with no details.

Figure 13: Horns of the Dall sheep (http://en.wikipedia.org/wiki/Dall_Sheep, 25/4/09)
It was not until D’Arcy Wentworth Thompson’s ‘On Growth and Form’, published in 1917, that the mechanics were addressed. Thompson (1992) argued that biologists were over emphasising evolution as the fundamental determinant of form and underemphasising the roles of the physical laws and mechanics:
Cell and tissue, shell and bone, leaf and flower, are so many portions of matter, and it is in obedience to the laws of physics that their particles have been moved, moulded and conformed.
(Thompson, 1992, p. 7)
He insisted that there were many forms in the natural world, such as the spiral horns of a Dall sheep, that were shaped not by evolution, but as a direct consequence of the conditions of growth or the forces in the environment’ (Ball, 2009, p. 12). ‘On growth and form’ was the first book to provide a formal analysis of pattern and form in nature. Yet Thompson’s morphological mapping only highlights, rather than addresses, the key underlying question: From where do these forms come from and how are they altered over evolutionary time? (Ball, 2009, p. 258).
Since the mid-1980s, discoveries are revealing how biological shape and form is indeed created. Advancements in developmental biology and evolutionary developmental biology, dubbed Evo Devo for short, are revealing a great deal about the invisible genes and some simple rules that shape animal form and evolution (Carroll, 2007, p. x). Carroll (2007, p. 283) claims that Evo Devo can be considered the third major act in a continuing evolutionary synthesis – illustrating how changes in development and genes are the basis of evolution.
In the 18th-century ‘it was assumed that creatures start life as miniature but fully formed versions of their adult selves, and just grow bigger’ (Ball, 2009, p. 260). This notion of ‘preformation’ in embryo development has been shown to be inaccurate. In 1859, Darwin’s proposed the idea that all forms are descended from common ancestors. Evo Devo has added significant weight to this argument through the advancements in embryogenesis which demonstrate a shared genetic tool kit used in embryo development.
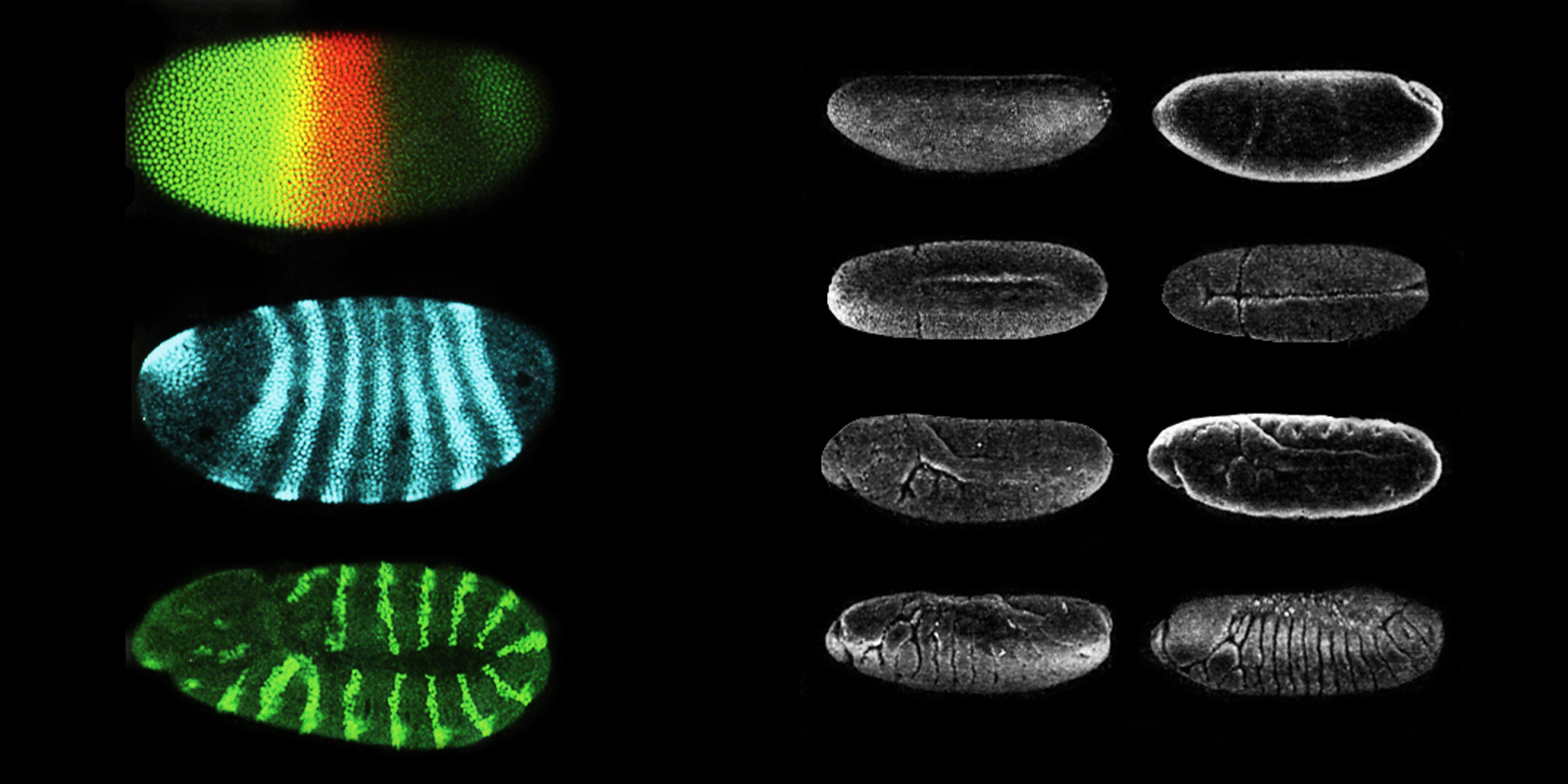
Figure 14 (Left): Genes in the fruit fly embryo are ‘switched on’ in stripe patterns in the early stages of growth (Carroll, 2007)
Figure 15 (Right): A fruit fly embryo develops grooves that become compartments of its segmented body (Carroll, 2007, p. 87)
Through the study of fruit fly Drosophila melanogaster, geneticists are showing that morphogenesis is a process of gradual elaboration. This is not a process of spontaneous, arbitrary patterning, but rather through careful subdivision via the so-called ‘homeotic’ gene. As Ball (2009, pp. 274-275) elaborates, when the DNA sequences of homeotic genes were examined for various mutations of fruit flies, it was found that all of them contained a short stretch of DNA that was identical. This segment was named ‘homeobox’ and the genes containing it were dubbed ‘Hox’ genes.
Hox genes have also been identified in humans and other mammals which imply that Hox genes are a rather ancient genetic element that determines body plans. By simply turning genetic dials and throwing genetic switches, the Hox genes control the organising logic of cellular differentiation from egg to adult (Ball, 2009, p. 271). However, it is important to note that these genes are not the drivers of evolution. The genetic tool kit represents possibility – realisation of its potential is ecologically driven (Carroll, 2007, p. 286).
The formation of architectural body plans

Figure 16: Greg Lynn’s Embryological House (Rocker, 2006, p. 92)
Genetic algorithms are grounded on Darwin’s notion of evolution. Yet, as it has been argued, Darwin’s theory put too much emphasis on evolution and not on where the forms came from or how they evolved. As morphogenetic architects are turning to biology for inspiration, could the use of genetic algorithms within architectural design be falling into the same trap? Poised with the new knowledge that biological morphogenesis is controlled by Hox genes, shouldn’t this logic be integrated into the genetic algorithm?
Although still in an embryological stage itself, the notion of Hox logic playing an integral role in genetic algorithms is starting to be explored by many. The philosopher Manuel de Landa has written extensively on the topic. De Landa (2002, p. 9) claims that in a sense, ‘evolutionary simulations replace design, since artists can use this software to breed new forms rather than specifically design them’. He argues that if the designer uses virtual evolution as a design tool to generate new forms and then uses this information to judge the aesthetic fitness of the outcomes, it transforms the role of the architect into ‘the equivalent of a prize-dog or a race breeder’ (DeLanda, 2002, p. 11).
Moreover, he claims that if evolved architectural structures are to enjoy the same degree of combinatorial productivity as biological ones they must begin with an adequate ‘abstract vertebrate’ (DeLanda, 2002, p. 12). By abstract vertebrate, de Landa is referring to the ‘body plan’ found in embryogenesis which shows the destinies of different regions of an embryo. De Landa is therefore arguing that breeding and evolution are not enough and that the architectural processes must include the logic homeotic genes if we are to progress into the enormous diversity of a post-Cambrian architectural period.

Figure 17: Bankside Paramorph by Decoi (http://web.mit.edu/mg_decoi/www/bankside/index02.html, 25/4/09)
Philip Steadman (2008) and Mark Goulthorpe (2008) have both explored the notion of a ‘Hox genetic algorithm’. Steadman has identified that one of the main issues with evolution within the genetic algorithm is that it requires initial forms from which breeding can occur. He proposes an alternative approach where a design might instead be ‘grown’ in some rather metaphorical sense from the germ or seed of an idea. The design process then becomes closer to the development of an individual from egg to adult rather than to the evolution of species. This he argues would be better suited to the ‘environment’ of the designer’s critical evaluations. Similarly, Goulthorpe (2008, pp. 149-150) promotes not the biological metaphor but the biological logic – the principle of differentiated speciation from restricted ancestry.
If a morphogenetic approach to the architectural design process is to succeed, the evolutionary architecture pioneered by architects such as John Frazer, must be further developed to encompass the genetic code at the most fundamental level. To date, architects have addressed this problem only discursively or in a very superficial way. In Greg Lynn’s Embryological House for instance, the ‘seed’ for the design is defined, then the computer generates all the mutations, producing a series of dwellings. But as de Landa has argued, the diversity is negligible compared to nature’s diversity. Similarity in Mark Goulthorpe’s Bankside Paramorph in London, none of the Hox logic that he is addressing in his essays is visible in his design.
So while many agree that the affordance offered by combining Hox logic with the genetic algorithm is enormous, it has yet to come to prominence within architectural circles. This may be attributed to the not-yet-calculated ‘Hox genetic algorithm’ with which scientists are still coming to terms. If this is indeed the case, architects must take charge and stop relying of biology to provide all the answers. The lack of appropriation of biological algorithms into architectural algorithms has stagnated the design process producing only limited possibilities. After all, architects must remember that they are creating models for something, not models of something.
Conclusion

Figure 18: Greg Lynn’s Embryological House (Rocker, 2006, p. 92)
This paper opened by claiming that architecture has frequently drawn inspiration from nature in its forms and structures. More recently however, a deeper understanding of nature’s beauty was being integrated into the architectural design process by an understanding of nature’s morphological processes. It was shown that the introduction of the genetic algorithm is fundamentally a response to Darwin’s notion of evolution. It was argued that just as Darwin’s theory of evolution was inadequate in describing where forms came from and how they evolved, so too is the genetic algorithm. Furthermore, the shift from form-making to form-evolving potentially limited the role of the designer to a kind of prize-dog breeder.
Inspired by the recent advancements in developmental biology and its ability to describe the mechanics of evolution, it was proposed that by modifying the genetic algorithm to encompass Hox logic, architecture can progress into a period of ‘post-Cambrian’ architectural diversity. It was argued that this process of ‘growing’ an idea, rather than evolving an idea, would be better suited to the architectural design process. In sum, the affordance offered by combining Hox logic with the genetic algorithm is enormous but its success lies in architects developing their own algorithm to address specifically their project. Although this may be perceived as somewhat overwhelming, architects can take respite in the knowledge that of all the possible rules that govern nature’s complexity, scientists have found that nature often uses the simplest.
Bibliography
Aranda, B., & Lasch, C. (2006). Tooling. New York: Princeton Architectural Press.
Ball, P. (2009). Nature’s patterns: A tapestry in three parts. Oxford: Oxford University Press.
Carroll, S. (2007). Endless forms most beautiful: the new science of Evo Devo and the making of the animal kingdom. London: Phoenix.
Darwin, C. (1859). The origin of species. London: John Murray.
DeLanda, M. (2002, Jan). Deleuze and the Use of the Genetic Algorithm in Architecture. Architectural Design: Contemporary Techniques in Architecture , pp. 9-12.
Flake, G. W. (1998). Genetics and evolution. In The computational beauty of nature (pp. 339-360). Cambridge: MIT Press.
Frazer, J. (1995). A natural model for architecture. In An evolutionary architecture (pp. 9-21). London: AA Press.
Gell-Mann, M. (1994). Information & crude complexity. In The quark and the jaguar (pp. 23-41). London: Abacus.
Gell-Mann, M. (1994). Machines that learn or simulate learning. In The quark and the jaguar (pp. 307-325). London: Abacus.
Giralt-Miracle, D. (2002). Gaudi: Exploring form. Barcelona: Lunwerg.
Goulthorpe, M. (2008). HOX aesthetics: The restrained profligacy of second-order generative processes. In B. Kolarevic, & K. Klinger (Eds.), Manufacturing material effects: Rethinking design and making in architecture (pp. 145-150). New York: Routledge.
Hersey, G., & Freedman, R. (1992). Possible Palladian Villas: (Plus a few instructively impossible ones). Cambridge: MIT Press.
Holland, J. (1996). Hidden order: How adaptation builds complexity. New York: Basic Books.
Leach, N. (2009, Jan/Feb). Digital morphogenesis. Architectural design: Theoretical meltdown , pp. 32-37.
Rocker, I. (2006, July/ Aug). Calculus-Based Form: An Interview with Greg Lynn. (H. Castle, Ed.) Architectural Design: Programming Cultures: Art & Architecture in the Age of Software , 76 (4), pp. 88-95.
Steadman, P. (2008). Afterword: Developments since 1980. In The evolution of designs: Biological analogy in architecture and the applied arts (pp. 237-270). London: Routledge.
Terzidis, K. (2006). Algorithmic architecture. Oxford: Elsevier.
Thompson, D. (1992). On growth and form. New York: Cambridge University Press.
Verebes, T. (Ed.). (2008). DRL ten : A design research compendium. London: AA Publications.




{kind=link}